
Lorsqu'on étudie la dynamique de croissance d'une population ou d'un organisme, on cherche à expliquer et éventuellement à prévoir l'évolution de celle-ci en fonction du temps. On propose alors un modèle, c'est-à-dire une représentation simplifiée sous forme mathématique de la dynamique observée. Une approche déterministe du problème consiste à établir une équation différentielle donnant une relation entre la fonction "taille" (de la population, de l'organisme) et sa dérivée dans le temps.
Selon Malthus (1798) [1], en l'absence de limitation par la quantité de ressources disponibles, toute population croîtrait selon une progression géométrique. Dans ce cas la croissance est exponentielle.
Considérons par exemple le nombre de cas cumulés de COVID-19 en France au moment du rebond épidémique ("deuxième vague") [2]. On étudie ici la population des français ayant contracté cette maladie.
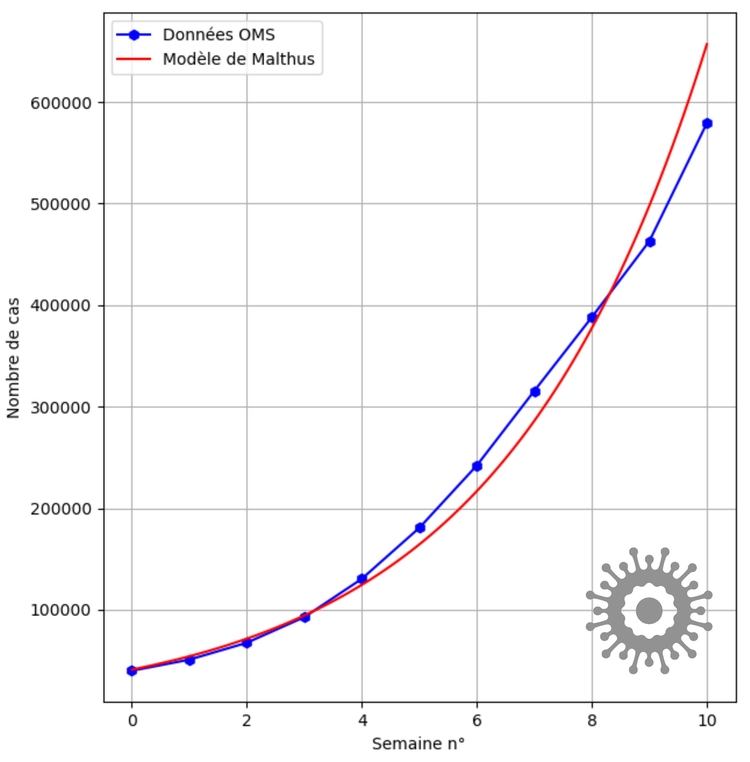
Les données disponibles correspondent assez bien à un modèle malthusien, autrement dit la taille de la population $N$ est solution de l'équation différentielle $N'=rN$, avec $r>0$ (sur la figure $r\simeq 0,277$).
Si on fait maintenant l'hypothèse que la croissance d'une population ou d'un organisme ralentit à partir d'un certain effectif du fait des contraintes du milieu, on considère cette fois des modèles de croissance dite "logistique" ou "régulée".
Considérons par exemple :
- le nombre d'éléphants du parc Kruger, un parc naturel protégé en Afrique du Sud créé au début des années 1900 dans l'espoir de permettre à une population d'éléphants quasiment éteinte à l'époque de croître naturellement [3] ;
- l'évolution de la masse corporelle d'un rat musqué au cours de sa vie [4].
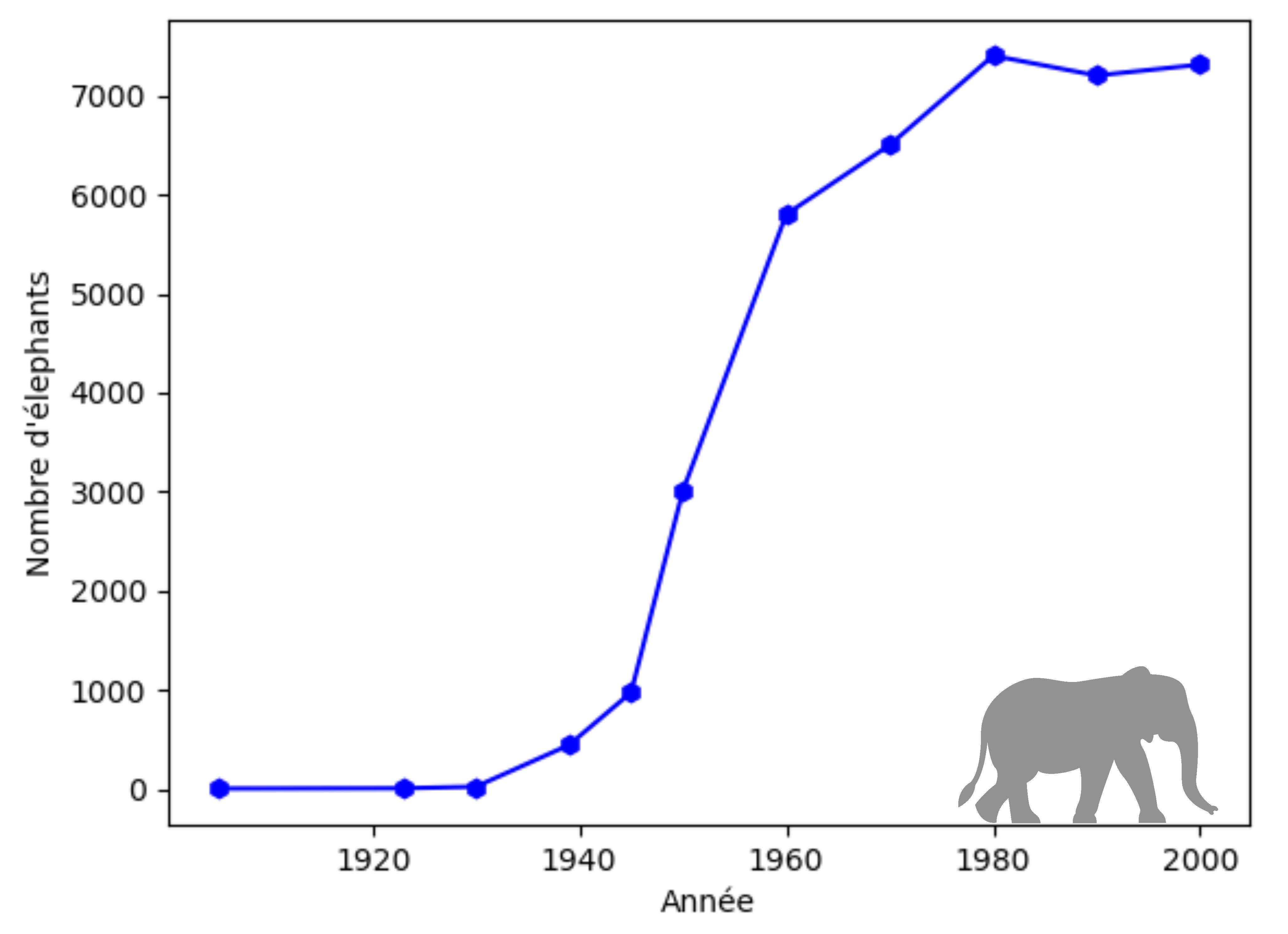
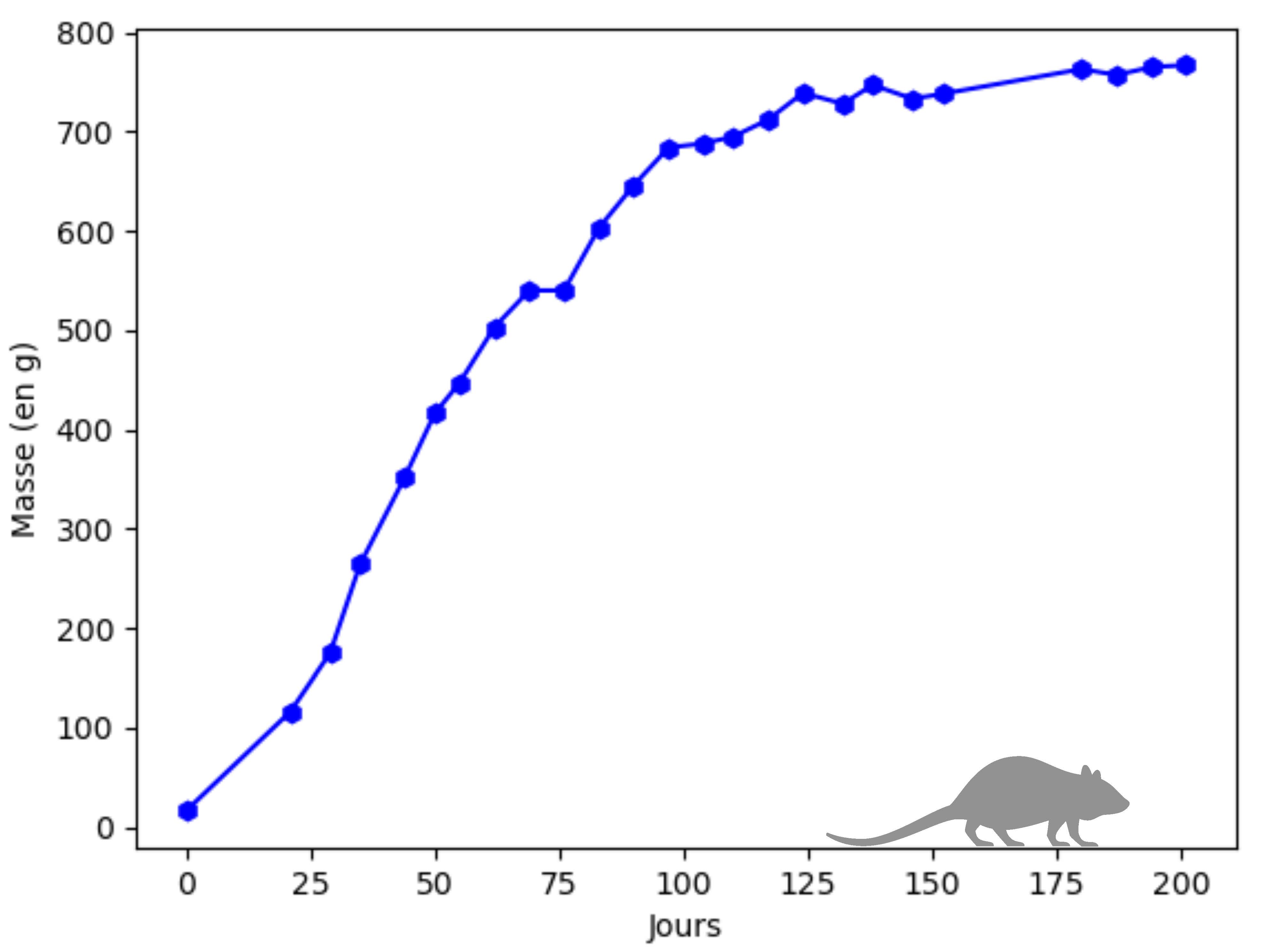
Dans les deux cas, il est clair qu'on observe un ralentissement de la croissance, ce qui se traduit par la présence d'un point d'inflexion sur la courbe. Le modèle de Malthus ne convient plus.
Les deux modèles logistiques les plus connus pour décrire ce type de croissance sont les modèles de Verhulst et de Gompertz. Ils permettent tous les deux de prendre en compte une capacité d'accueil du milieu limitée. Ces modèles s'attachent principalement à décrire la dynamique de la courbe de croissance sans chercher une explication biologique du phénomène de ralentissement de la croissance.
Dans les deux cas, on résout une équation différentielle de la forme $N'=f(N)$ où la fonction $f$ décrit l'évolution de la croissance de la population en fonction de la progression de celle-ci.
On va présenter ces deux modèles, déterminer explicitement les solutions correspondantes, identifier leurs points communs et leurs différences notables pour enfin construire un nouveau modèle de croissance logistique permettant notamment d'ajuster la position du point d'inflexion de la courbe représentant une solution.
Deux modèles historiques de croissance logistique
Le modèle de Verhulst
Verhulst a proposé en 1838 de modifier le modèle de croissance exponentielle de Malthus afin de modérer la croissance de la population étudiée en prenant en compte les ressources disponibles. En notant $N$ le nombre d'individus de la population étudiée, le modèle de Verhulst affirme que :
$$\forall t\in\mathbb{R}_+, \qquad N'(t) = rN(t)\left(1-\frac{N(t)}{K}\right), \qquad (V)$$
où $r$ est le coefficient de croissance intrinsèque et $K$ la capacité d'accueil du milieu.
En utilisant ce modèle, on considère que si $N\in]0;K[$, alors $N'>0$ ce qui traduit une croissance de la population1. Pour des petites populations, l'accroissement est proportionnel au nombre d'individus (car alors $1-\frac{N(t)}{K} \simeq 1$) donc la croissance est exponentielle dans un premier temps, mais l'accroissement atteint ensuite une valeur maximale à partir de laquelle la croissance ralentit et la population tend vers la capacité d'accueil du milieu $K$.
Cette évolution est cohérente avec celle de la population d'éléphants au parc Kruger pour laquelle on identifie sur la figure 2 que $K\simeq 7500$ ou avec celle de la masse corporelle du rat musqué pour laquelle on identifie sur la figure 3 que $K\simeq 800$.
Pour $0<N(0)<K$, la solution de l'équation différentielle $(V)$ associée à la condition initiale $N(0)=N_0$ est :
$$N : t \mapsto \dfrac{K}{1+e^{-(rt+C_0)}},\quad \text{avec }C_0=\ln\left( \dfrac{N_0}{K-N_0} \right).$$
Notons $N$ l'unique solution au problème de Cauchy constitué par l'équation $(V)$ et la condition initiale $N(0)=N_0$. Commençons par noter que si $0<N_0<K$, alors pour tout $t\in\mathbb{R}_+$, $0<N(t)<K$. En effet,
- si $N$ s'annule en $t_0$, par unicité de la solution au problème de Cauchy constitué par l'équation $(V)$ et la condition initiale $N(t_0)=0$, $N$ est la fonction nulle ce qui est exclu ;
- si $N$ prend la valeur $K$ en $t_1$, par unicité de la solution au problème de Cauchy constitué par l'équation $(V)$ et la condition initiale $N(t_1)=K$, $N$ est la fonction constante égale à $K$ ce qui est exclu.
Comme $N$ ne s'annule pas, on a que :
$$N' = rN\left(1-\dfrac{N}{K}\right)\iff N' = rN-\dfrac{r}{K}N^2 \iff \underbrace{-\dfrac{N'}{N^2}}_{=y'} = -r\underbrace{\dfrac{1}{N}}_{=y}+\dfrac{r}{K}$$
donc $y=\dfrac{1}{N}$ est solution de l'équation différentielle $y'+ry = \dfrac{r}{K}$. La fonction $y$ est solution d'une équation différentielle linéaire d'ordre 1 à coefficients et second membre constants.
On trouve alors qu'il existe une constante $C\in\mathbb{R}$ telle que : $y : t \mapsto Ce^{-rt}+\dfrac{1}{K}$
Puis $N = \dfrac{1}{y}$ donc : $$\forall t\geq 0, \quad N(t)=\dfrac{1}{Ce^{-rt}+\dfrac{1}{K}}=\dfrac{K}{CKe^{-rt}+1}$$
Enfin, $$N(0)=N_0 \iff \dfrac{K}{CK+1}=N_0 \iff C = \dfrac{1}{N_0}-\dfrac{1}{K}$$
d'où la solution suivante au problème avec condition initiale : $$N : t \mapsto \dfrac{K}{\left(\frac{1}{N_0}-\frac{1}{K}\right)Ke^{-rt}+1}=\dfrac{K}{\frac{K-N_0}{N_0}e^{-rt}+1}=\dfrac{K}{e^{-\ln\left(\frac{N_0}{K-N_0}\right)}e^{-rt}+1}=\dfrac{K}{e^{-(rt+C_0)}+1}$$
Il est tout à fait remarquable qu'il soit possible d'exhiber la solution exacte de l'équation $(V)$ associée à la condition initiale $N(0)=N_0$ puisque l'équation différentielle $(V)$ n'est pas linéaire.
Notons enfin qu'en posant le ratio $x=\dfrac{N}{K}$, l'équation devient $x' = rx (1-x)$ et la solution associée à la condition initiale $x_0=\dfrac{N_0}{K}$ est : $$x : t \mapsto \dfrac{1}{1+e^{-(rt+\tilde{C}_0)}} \quad \text{où }\tilde{C}_0 = \ln\left( \dfrac{x_0}{1-x_0} \right).$$
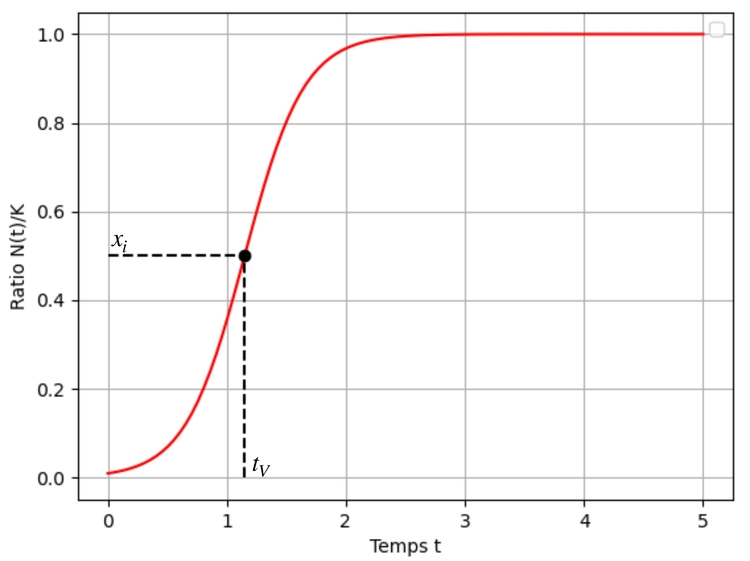
Identifions les coordonnées du point d'inflexion : pour une fonction $x$ deux fois dérivable, si la vitesse de croissance $x'$ ralentit alors l'accélération $x''$ s'annule. Or ici : $$x'' = 0 \iff r(1-2x)=0 \iff x=\dfrac{1}{2}$$
autrement dit l'ordonnée du point d'inflexion est $x_i=\dfrac12$.
L'abscisse $t_{V}$ correspondante est solution de l'équation $x(t_V) = \dfrac12$. Or : $$x(t_V) = \dfrac12 \iff t_V = -\dfrac{\tilde{C}_0}{r}$$
On retient surtout à ce stade qu'en utilisant ce modèle, la croissance commence à ralentir lorsque la population atteint la moitié de la capacité d'accueil $K$ du milieu. Cela correspond assez bien à ce qu'on observe dans le cas de l'évolution de la population d'éléphants au parc Kruger.
Le modèle de Gompertz
Le modèle de Gompertz est un modèle concurrent du modèle de Verhulst pour la modélisation de la croissance des organismes. Il est notamment utilisé pour ajuster les données de la croissance des tumeurs. Les tumeurs peuvent en effet être considérées comme des populations de cellules dans un espace restreint, où la disponibilité en nutriments est limitée. En notant $N$ le nombre de cellules cancéreuses et $K$ la capacité limite du milieu, le modèle de Gompertz affirme que :
$$\forall t\in\mathbb{R}_+, \qquad N'(t) = aN\ln\left(\dfrac{K}{N}\right). \qquad (G)$$
Ce modèle s'est avéré en pratique très performant pour étudier la croissance bactérienne comme celle des tumeurs. De plus, on peut comme pour le modèle précédent calculer la solution exacte de cette équation différentielle, ce qui explique en partie son franc succès dans le monde médical.
La solution de l'équation différentielle $(G)$ associée à la condition initiale $N(0)=N_0$ est : $$N : t \mapsto Ke^{be^{-at}},\quad \text{avec }b=\ln\left(\frac{N_0}{K}\right).$$
Notons $N$ l'unique solution au problème de Cauchy constitué par l'équation $(G)$ et la condition initiale $N(0)=N_0$. Commençons par noter que si $0<N_0<K$, alors pour tout $t\in\mathbb{R}_+$, $0<N(t)<K$ pour la même raison que vue précédemment avec l'équation $(V)$. Comme $N$ est une fonction strictement positive, on a que :
$$N' = aN\ln\left(\dfrac{K}{N}\right) \iff \underbrace{\dfrac{N'}{N}}_{=y'} = a\ln(K)-a\underbrace{\ln(N)}_{=y}$$
donc $y=\ln(N)$ est solution de l'équation différentielle $y'+ay = a\ln(K)$.
La fonction $y$ est solution d'une équation différentielle linéaire d'ordre 1 à coefficients et second membre constants. On trouve alors qu'il existe une constante $b\in\mathbb{R}$ telle que : $$y : t \mapsto be^{-at}+\ln(K)$$
Puis $N = e^y$ donc : $$\forall t\geq 0, \quad N(t)=e^{be^{-at}+\ln(K)}=Ke^{be^{-at}}$$
Enfin, $$N(0)=N_0 \iff Ke^{b}=N_0 \iff e^b = \dfrac{N_0}{K} \iff b = \ln\left( \dfrac{N_0}{K} \right)$$
d'où la solution suivante au problème avec condition initiale : $$N : t \mapsto Ke^{be^{-at}} \text{ avec } b =\ln\left(\frac{N_0}{K}\right)$$
En posant le ratio $x=\dfrac{N}{K}$, l'équation devient $x' = -ax \ln(x)$ et la solution associée à la condition initiale $x_0=\dfrac{N_0}{K}$ est : $$x : t \mapsto e^{\tilde{b}e^{-at}} \quad \text{où }\tilde{b} = \ln\left( x_0\right).$$
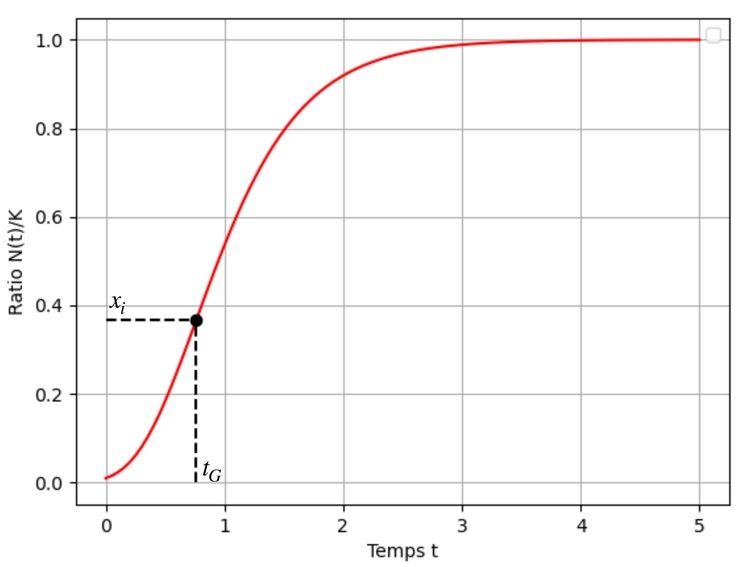
Identifions les coordonnées du point d'inflexion : de nouveau, si la vitesse de croissance $x'$ ralentit alors l'accélération $x''$ s'annule. Or ici : $$x''=0 \iff -a(\ln(x)+1)=0 \iff x=\dfrac{1}{e}$$
autrement dit l'ordonnée du point d'inflexion est $x_i=\dfrac{1}{e}\simeq 0,37$.
L'abscisse $t_{G}$ correspondante est solution de l'équation $x(t_G) = \dfrac{1}{e}$. Or : $$x(t_G) = \dfrac{1}{e} \iff t_G = \dfrac{\ln\left(\ln\left(\dfrac{1}{x_0}\right) \right)}{a}$$
On retient cette fois qu'en utilisant ce modèle, la croissance commence à ralentir lorsque la population atteint $\dfrac{K}{e}$ à savoir strictement moins que la moitié de la capacité du milieu. Cela correspond assez bien à ce qu'on observe dans le cas de l'évolution de la masse corporelle d'un rat musqué.
Une inflexion différente mais imposée
Les modèles de Verhulst et de Gompertz donnent bien des évolutions de population similaires. La différence fondamentale réside dans la position du point d'inflexion de la courbe : à $K$ fixé, la population au moment où la croissance commence à ralentir du fait de la limitation des ressources n'est pas la même mais est imposé par le modèle choisi. Ni l'un ni l'autre de ces deux modèles ne permet d'obtenir une inflexion au tiers de la capacité $K$ du milieu par exemple!
Identification des points communs entre les courbes de croissance
Bien qu'il y ait des différences entre les deux modèles, les fonctions : $$f_V : x \mapsto rx(1-x) \quad \text{et} \quad f_G : x \mapsto -ax\ln(x)$$
autrement dit les fonctions décrivant la vitesse de croissance de la population, ont une allure similaire.
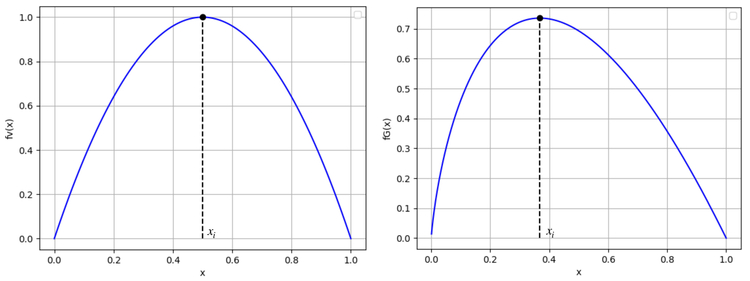
On retrouve ici l'observation formulée dans le paragraphe précédent : les dérivées des fonctions $f_V$ et $f_G$ s'annulent respectivement en $x_i=\frac12$ et $x_i=\frac{1}{e}$.
Quels sont les points communs entre ces deux courbes de croissance?
- il s'agit dans les deux cas de fonctions d'abord croissante puis décroissante sur $]0;1[$;
- $f_V(0)=0=\displaystyle\lim_{x\to 0^+}f_G(x)$;
- $f_V(1)=0=f_G(1)$.
Il est alors raisonnable de penser que si $f$ est une fonction respectant ces trois contraintes, l'équation $x'=f(x)$ assortie de la condition initiale $x_0\in]0;1[$ aura pour solution une fonction logistique dont l'allure sera sensiblement similaire aux courbes présentées dans les deux premières parties. Autrement dit, en se concentrant sur l'allure de la courbe de croissance, on peut créer un nouveau modèle logistique original! Si on arrive de surcroit à imposer que la dérivée de la fonction de croissance s'annule en une quantité $x_i\in]0;1[$ imposée, cela signifie qu'on pourra choisir notre modèle en fonction de l'ordonnée du point d'inflexion désiré.
Construction de nouveaux modèles de croissance logistique
Une fonction de croissance polynomiale de degré $3$
Considérons la fonction $f:x\mapsto x(1-x)^2$. Cette fonction est dérivable sur $[0;1]$ de dérivée $f'(x)=(1-x)(1-3x)$ ce qui nous permet d'obtenir l'allure suivante:
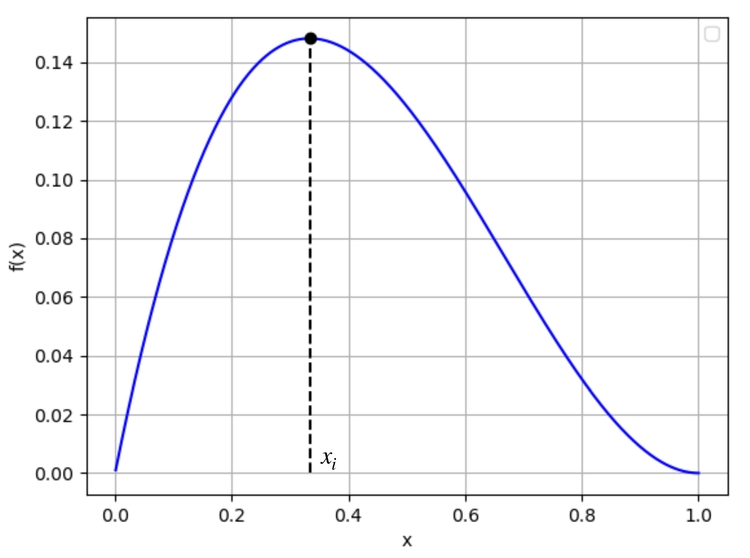
En résolvant $x'=f(x)$, on va donc obtenir une courbe logistique avec une inflexion pour $x_i=\frac13$, puisque c'est la valeur en laquelle $f'$ s'annule.
Une résolution numérique du problème de Cauchy $x'=f(x)$ et $x(0)=x_0$ donne le graphe de $x$ suivant :
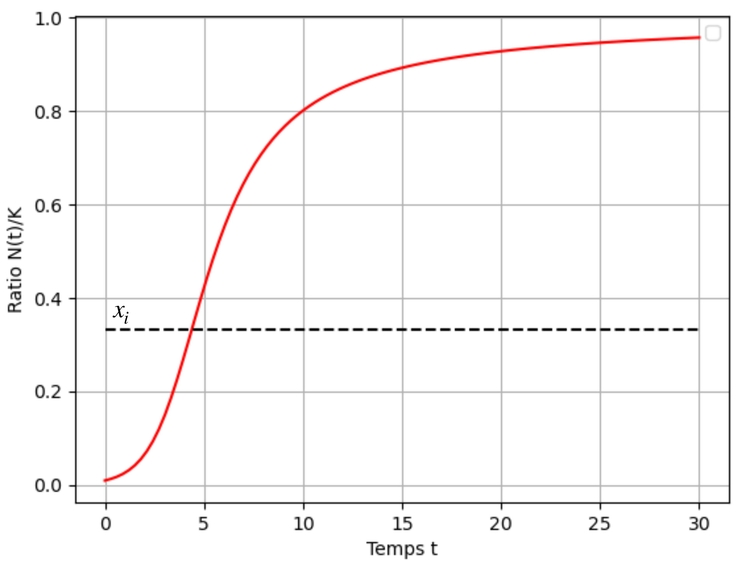
Remarquons cependant que la courbe de la fonction $f$ choisie admet une tangente horizontale au point d'abscisse $1$ ce qui a pour conséquence de ralentir la convergence du ratio $x$ vers $1$. Cette particularité n'était pas présente sur les courbes des fonctions de croissance des modèles de Verhulst et Gompertz.
Une famille de fonctions de croissance permettant d'ajuster la position du point d'inflexion
Soit $\alpha\in]-1;+\infty[\setminus${0,1}. Considérons la fonction $g_\alpha:x\mapsto x(1-x^\alpha)$. Cette fonction est dérivable sur $]0;1]$ de dérivée $g_\alpha'(x)=1-(1+\alpha)x^\alpha$ ce qui nous permet d'obtenir l'allure suivante :
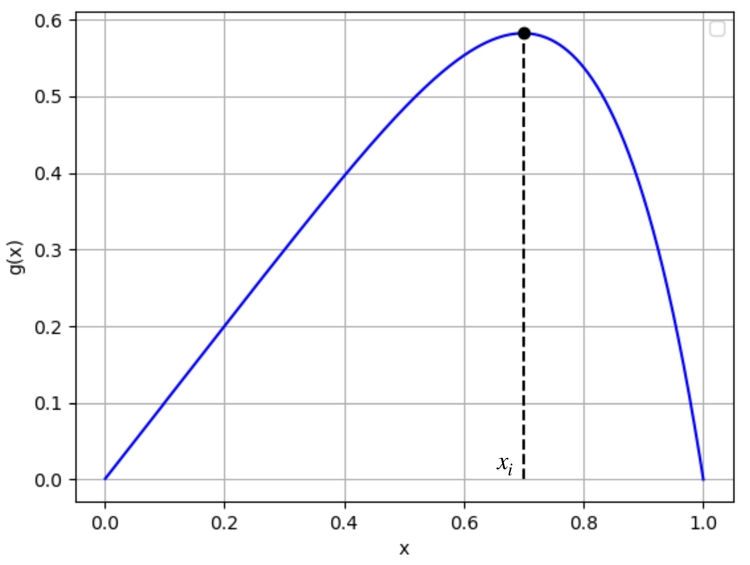
On trouvera en résolvant $x'=g(x)$ une courbe logistique avec un point d'inflexion d'ordonnée $x_i=\left(\frac{1}{1+\alpha}\right)^{\frac1{\alpha}}$ ($\simeq 0,7$ si $\alpha=5$), puisque c'est la valeur en laquelle $g'$ s'annule.
Une résolution numérique du problème de Cauchy $x'=g_\alpha(x)$ et $x(0)=x_0\in]0;1[$ donne l'évolution de population suivante :
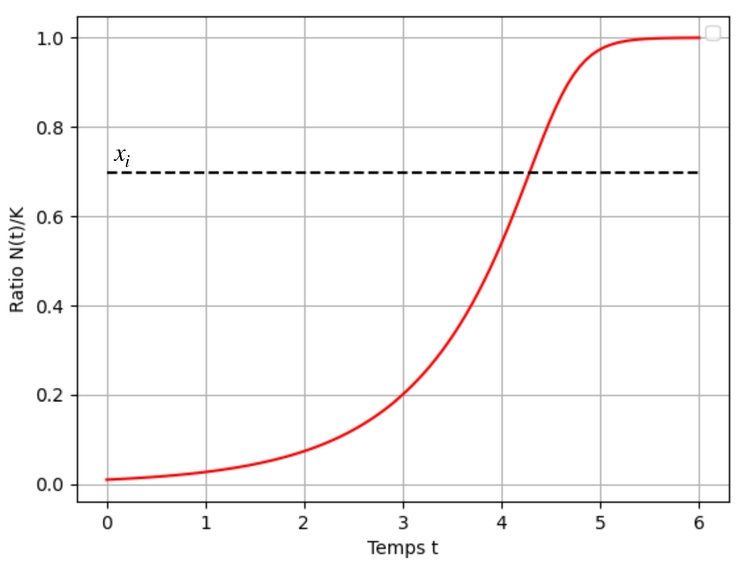
Avec ce deuxième exemple, on obtient une famille de fonctions permettant d'ajuster l'inflexion de la solution à une proportion souhaitée de la capacité du milieu! En effet, une rapide étude de la fonction $u : \alpha \mapsto \left( \dfrac{1}{1+\alpha}\right)^{\frac{1}{\alpha}}$ permet de justifier qu'elle réalise une bijection de $]-1;+\infty[$ sur $]0;1[$. Autrement dit quelque soit la proportion $x_i$ de la capacité d'accueil $K$ à laquelle on souhaite que la vitesse de croissance ralentisse, il existe un unique réel $\alpha\in]-1;+\infty[$ tel que $u(\alpha)=x_i$. En résolvant l'équation $x'=g_\alpha(x)$ correspondant à ce réel $\alpha$, on obtient une solution dont la courbe comporte une inflexion au point d'ordonnée $x_i$.
Conclusion
On a ainsi créé une famille de fonctions de croissance permettant d'obtenir des courbes logistiques dont le point d'inflexion peut être situé à quelle que proportion $x_i$ que ce soit de la capacité d'accueil $K$ du milieu. Elles ont cependant un vilain défaut en comparaison des populaires modèles de Verhulst et Gompertz : il apparait cette fois bien plus ardu de trouver une solution explicite de l'équation différentielle non linéaire $x'=f(x)$. La courbe d'une solution de cette équation différentielle a été obtenue par une résolution numérique approchée. La recherche du temps $t_i$ auquel l'inflexion a lieu, autrement dit la résolution de l'équation $x(t) = x_i$, ne peut se faire que numériquement également.
Il est enfin possible d'ajuster l'expression de la fonction de croissance de manière à pouvoir choisir à la fois l'ordonnée $x_i$ du point d'inflexion de la courbe d'évolution mais aussi l'abscisse $t^*$ de ce point d'inflexion ! Il suffit pour cela de multiplier la fonction $f$ de croissance par $ \dfrac{t_i}{t^*}$.
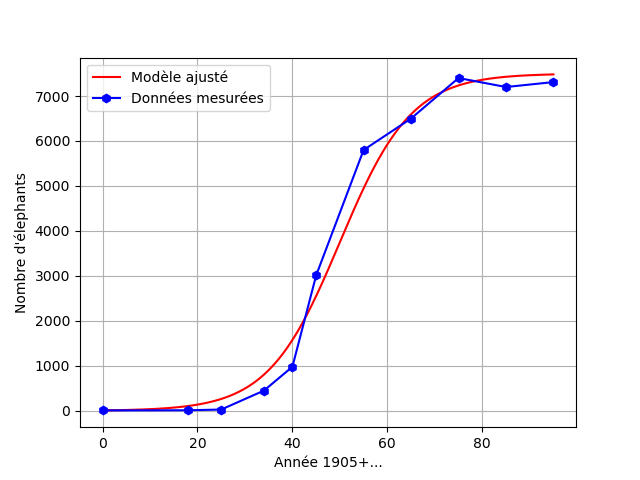
Par exemple, pour obtenir une inflexion en l'année $1955$ (estimation par lecture graphique) en utilisant le modèle de Verhulst précédemment étudié pour décrire la population d'éléphants du parc Kruger, on résout l'équation : $$N' = \dfrac{t_V}{t^*} \times r N\left(1-\dfrac{N}{K}\right)$$
et on obtient la solution suivante, ajustée aux données mesurées :
Références
- Thomas-Robert MALTHUS, Essai sur le principe de population, 1798
- Données disponibles le 14 janvier 2025 sur le site World Health Organization Data, https://data.who.int/dashboards/covid19/data
- Données disponibles le 14 janvier 2025 dans un cours de Mathématiques pour la biologie dispensé par l'Université de NiceTechno-Science, https://math.univ-cotedazur.fr/~diener/MAB0809/LOGISTI.pdf
- Données disponibles le 14 janvier 2025 sur le site Techno-Science, https://www.techno-science.net/glossaire-definition/Modele-de-Gompertz.html
- Pierre-François VERHULST, Notice sur la loi que la population suit dans son accroissement, 1838
- Benjamin GOMPERTZ, On the nature of the function expressive of the law of human mortality, and on a new mode of determining the value of life contingencies, 1833
Dans le programme de l'option mathématiques complémentaires (Terminale), les modèles de Malthus et de Verhulst sont cités comme problèmes possibles au sein du thème "Modèles d'évolution".


